La vie des plasmes
Le ribomonde (Ou l'ARN et les peptides prennent le relais)
Le ribomonde
Nous en somme au stade ou des particules d'origine interstellaire sont recouvertes d'une couche moléculaire comprenant des acides nucléiques liés à des peptides, flottent dans l'océan, se déposent sur le fond et les rives, environnés de gouttelettes sphéroïdales, parfois liés à elles. Les divers rayonnements subis, l'alternance de cycle de dessiccation et de ré-hydratation, même partielle, sont à l'origine d'un apport continuel d'énergie permettant aux synthèses de sa poursuivre. Nos paleoribosomes, attachés à des membranes primitives, voient leur complexité et leur taille s'accroitre. Ils contiennent des peptides de plus en plus long, de plus en plus repliés et complexes. Les acides nucléiques deviennent, à force de complexification, capable d'auto-reproduction. Qu'est ce qui les pousse vers la complexité? tout simplementl'obligation qu'ils ont de devoir résister à l'hydrolyse (à l'origine d'une véritable pression de sélection), et pour cela de posséder une séquence leur permettant de se replier de façon correcte (Eigen & al., 1981). Comme ce processus moléculaire est plus rapide que la croissance cristalline, les riboparticules qui contiennent ces séquences se "reproduisent", croissent de plus en plus vite. Leur masse augmentant, elles ont tendance à s'enfoncer dans le milieu liquide, générant ainsi des tractions de plus en plus grandes sur les membranes liées aux peptides, et jouant peut être un rôle de "flotteur". Ces forces sont à l'origine du repliement des membranes. Nos agrégats se dupliquent au niveau nucléique, mais pas au niveau protéique (qui ne sont pas autoreplicante). Il en résulte que seules les séquences autoreplicantes capables, par leurs conformations, de diriger des synthèses protéiques, sont favorisées: c'est l'origine du transfert d'information entre acides nucléiques et protéines, le début du code génétique.
Les peptides
liés aux acides
nucléiques, a force de se replier, acquièrent des
formes qui facilitent les réactions chimiques: ce sont
les premier proto-enzymes. Et sur quelles molécules vont
ils agir ? Celles qui leur sont, géographiques, les plus
proches, c'est à dire les acides nucléiques. Dès
cet instant, la collaboration entre enzyme et acides nucléique
prend naissance et ne cessera de se complexifier. Une des traces
actuelles de cette collaboration est que beaucoup de coenzymes sont des
nucléotides ou des dérivés de nucléotides
(comme le célèbre NAD (nicotinamide
adenine di-nucléotide) si utile aux
dosages enzymatiques!)
Les séquences d'acides nucléiques sont sujettes à mutations, à variations: une compétition se créée, et on obtient rapidement plusieurs quasi-espéces d'ARN. Cette évolution moléculaire a pu être obtenue in vitro (Eigen & al., 1981, Fontana, 1998), et avec des résultats extrêmement intéressant, montrant qu'un système moléculaire autoreproductif possède une adaptabilité proprement sidérante aux variations de son milieu (ce qui était indispensable dans l'environnement des débuts de la vie!). La reproductibilité des premiers acides nucléiques dépend de l'appariement de bases complémentaires, peut être aidé dès le début par des peptides de forme bien définie. Cette aide peptidique a permis de dépasser le seuil de 100 nucléotides au delà duquel les erreurs d'appariement ne permettent plus la conservation d'une séquence (qui devait être riche en G et C). Le système s'organise rapidement selon trois composantes :
Il doit subsister des traces de ces nombreux ARN différents. Actuellement, les chercheurs découvrent de nombreux types d'ARN (Bachellerie, 2003) dans les cellules, aux rôles plus variès que l'on ne l'imaginait. Ainsi, les ARN "interférents" sont impliqués dans le contrôle de l'expression des gènes. Dans le processus de maturation de ces ARN, une enzyme, la DICER, transforme une "épingle à cheveux" d'ARN en séquence double brin (Lau & Bartel, 2003). Ce processus a dû débuter avant même qu'il existe des cellules, et cette enzyme doit être très conservée chez tous les êtres vivants. C'est apparemment le cas.
Les biologiste moléculaires découvrent que le monde des ARN est bien plus complexe qu'on ne l'aurait cru. Ces molécules ubiquitaires jouent des rôles variés dans les cellules. Le tableau suivant, où les ARN sont classés par dimension (longueur) croissante, en donne une idée.
|
|
|
|
|
|
|
Se lie aux ARNm de certains gènes et les inactivent. Participent au controle de l'expression des gènes. Se lient fortement à l'ADN et changent sa forme. Sont synthétisés à partir d'"épingles à cheveux" d'ARN dont une enzyme, la DICER, coupe la "boucle". On obtient ainsi des ARN doubles brin aux extrémités "libres", qui peuvent éventuellement s'ouvrir, voire se séparer afin de se fixer sur d'autres molécules. Bloquent la traduction d'un ARNm |
|
|
ARN guides | Eliminent des nucléotides d'autres ARN en provoquant par appariement le repliement puis l'excision de certains nucléotides. |
|
|
ARN nucléolaires | Sert de point de fixation spécifique à des protéines qui trasnforment l'Uracyle en pseudo-urifdines |
|
|
ARN nucléaires des complexes d'épissage | |
|
|
ARN 7SL | Clef de reconnaissance des protéines en cours de synthèse, qui les oriente vers une des "citernes " de la cellule. |
|
|
ARN 6S | |
|
|
ARN transfert-messager | |
|
|
ARN télomérique | systéme de fixation d'enzymes variés permettant la synthèse des télomères en prévision des divisions cellulaires |
|
|
ARN XIST | |
| ARN messager | ||
| ARN de transfert | ||
C'est à ce niveau, avant même une séquestration nette dans des cellules séparées, que c'est élaboré le code génétique commun (à 99,9 %) à tous les organismes que nous connaissons. Néanmoins, on peut déjà dire qu'apparemment le code génétique s'est dès l'origine organisé en triplets de bases correspondante à un acide aminé. Cette organisation en triplet résulte peut être d'un problème de topologie, le repliement de l'ARN "de réserve" n'exposant que des suites de trois bases accessibles aux ARN " de transfert".
L'ancienneté des ARN de transfert est confirmée par leur structure très voisine chez des organismes pourtant très éloignés dans le temps et le champ de l'évolution:
 |
 |
 |
|
|
|
|
Il est extrémement intéressant que
les résultats de l'étude détaillée des
ribosomes actuels (Steitz, 2000) aient montré que
c'est bien
un ARN ribosomal, stabilisé, contraint par les
protéines ribosomiales, qui catalyse la principale
réaction chimique de liaison des acides aminés, à
savoir une action peptidyl transferase. Ceci démontre que,
conformément à ce que nous avons vu, le ribosome est
bien le reliquat actuel des premières formes de vies, le lieu
où c'est noué l'indispensable alliance des
protéines et des acides nucléiques.
 Structure d'un ribosome d'après les travaux de l'équipe de Steitz. Le site actif doit sa forme à l'assemblage protéine + AN mais son activité chimique est due aux ARN présents, se comportant comme un "ribozyme" et catalysant la polymérisation des acides aminés. Lien ci dessous vers ce remarquable laboratoire de l'université de Yale. |
 |
Le code
génétique, un interessant fossile moléculaire
- les levures candida décodent CUG comme donnant une serine (à la place de la leucine)
- les mycoplasmes et les mitochondries utilisent UGA comme codon du tryptophane, alors que "normalement", c'est un codon STOP. Le codon AUA correspond également chez ces dernières à la méthionine au lieu de l'isoleucine. Il existe même des variations du code entre mitochondries appartenant à des espèces différentes.
- chez les paramécies, UAG donne la glutamine et non STOP.
- les algues unicellulaires acetabularia traduisent les codons STOP en glycine
Ces variations se retrouvant dans diverses lignées évolutives, ont doit en conclure que le code génétique est le résultat d'une évolution s'étant déroulée pendant plusieurs milliards d'années durant lesquelles des codes concurrents se sont affrontés pour la domination du vivant. Les possibilités étaient, au départ, immenses: il existe environ 2x10(18) codes différents possibles !
Apparemment, ce code s'est dès
l'origine organisé en triplets de bases (appelés
"codon") correspondantes à un acide aminé.
Cette organisation en triplets résulte peut-être
d'un problème de topologie, le repliement de l'ARN "de
réserve" n'exposant que des suites de trois bases
accessibles aux ARNt. Cependant, l'examen
du code montre que ce sont surtout
les deux premières bases qui sont importantes pour
déterminer
un acide aminé (ci
dessous: code génétique
primitif avec triplet à deux bases codantes)
Un code à deux "lettres
codantes" aurait permis de distinguer 16 types d'acides aminés.
C'est largement plus qu'il n'en faut, car de nombreux acides
aminés peuvent être obtenus par modifications secondaire
du squelette carboné d'un autre acide aminé. Ainsi, la
glycine, le plus simple des acides aminés, peut il conduire
à une douzaine d'autres AA! Il est plus que probable qu'à
l'origine, le code génétique ne correspondanit
qu'à quelques acides aminés (de nos jours, quelques
organismes utilisent plus de 20 acides aminés en modifiant
chimiquement un des 20 AA classique alors même qu'il est
liè à son ARNt...). Peut ont faire des suppositions
sur le nombre initial d'AA codés? En se basant sur la nature
chimique des acides aminés, on peut se limiter à un
minimum de 3 AA originels (glycine, lysine, glutamine).
Les ancêtres des ARN de transfert auraient très bien pu se lier à davantage de bases de l'ARN, mais seules 2 de ces dernières auraient été déterminantes, la troisième le devenant de façon secondaire. On aurait très bien pu avoir aussi des ARNt ancestraux se liant à 7 bases de l'ARNm, par exemple, mais ne "lisant" que 3 de ces dernières.
Quelles sont les traces de l'ancienne conformation du code génétique ?
La topologie des chaînes
latérales
des acides aminés ainsi que le champ de forces qui leur
est associé présentent des similarités avec
celle de leur codon. Dans certains cas, le codon serait capable
de se mouler sur les chaînes latérales. De plus,
Certains acides aminés seraient capables de s'intercaler
entre les bases de leur codon, se disposant de façon
perpendiculaire
à la chaîne d'ARN. Ce sont là des indices
montrant qu'à l'origine il y avait bien un lien direct
entre ARN et monomères des protéines: le code
génétique
ne résulte pas d'un processus aléatoire:

Les premiers ARN
à s'être
liés aux acides aminés ont constitué les
ancêtres des ARN de transfert: leur ancienneté est, nous
l'avons vu, confirmée par leur forme très
voisine chez des organismes très éloignés
évolutivement.
Code génétique
de "toutes" les formes de vie terrestre connues actuelles --->
L'universalité du code génétique limite fortement le nombre d'ancêtres communs: ce code commun nous indique qu'il n'y a apparemment qu'une quasi-espéce moléculaire qui a réussi à prendre l'ascendant sur toutes les autres. Pourquoi? La question reste ouverte, et l'on peut invoquer par exemple une meilleure disponibilité des acides aminés du code, ou une efficacité supérieure des synthèses qu'il dirige. Des recherches de simulation informatique de codes alternatifs (Freeland & Hurst, 1998, 2004) ont identifiées une caractéristique supplémentaire du code mettant en lumière les pressions de sélection auxquelles il a été soumis : le code actuel se révèle posséder 2 caractéristiques principales: il minimise les risques d'erreurs critiques (causées par un mauvais repliement des protéines) en exerçant une forte contrainte sur le caractère hydrophyle ou hydrophobe des AA alors qu'il "permet" des erreurs légères qui sont une des forces motrices de l'évolution des organismes.
l’étude du code génétique par modélisation informatique (Ycas M, 1999)
montre que ce dernier ne correspond pas à un optimum de
résistance aux erreurs de lectures et aux mutations. Des
relations existant entre codon et propriétés physiques
des AA, on en déduit que le
lien codon-AA a été établit dès la mise en
place de l’ancêtre de l’aminoacyl synthetase qui a
lié AA et tRNA, avant que le systéme actuel
ne soit en place. On retrouve ici la préexistence de
l'ancêtre des ribosomes par rapport au code
génétique, renforçant mon idée selon
laquelle les ribosomes sont les reliquats des premières formes
de vie.
Le code génétique est redondant, plusieurs triplets
correspondant à un même acide aminé. Cette
redondance pourrait s'expliquer par la coexistence ancienne de
plusieurs codes différents, provenant de formes de vie
différentes, qui se seraient par la suite organisés
en un ensemble unique, par transfert d'information résultant
de processus de fusion de cellules, à l'occasion des premiers
phénomènes de nutrition par exemple, ou du
développement
de parasites ou de virus. Le tout était de parvenir à
un échange de différents ARN de transfert, dont
les séquences furent par la suite incorporées dans
le génome ADN, lorsque cette molécule fit son apparition.
Séquestration
Le problème des membranes protectrices
est d'une grande importance pour l'origine de la vie.Si l'on se
rappelle la fragilité des polymères biologiques
vis à vis de l'eau, le sel moyen de les protéger
d'un dilution fatale est de les envelopper dans une membrane fortement
hydrophobe.
Dans les cellules modernes, les membranes sont constituées de protéines insérées dans une double couche de phospholipides. La synthèse de ces dernier réclame des enzymes spécialisés, et on ne peut donc supposer qu'ils ait été présents à l'aube de la vie. D'autres composés présentant des propriétés similaires devaient assurer leur fonction. Le point commun à des molécules devant former des vésicules au contact de l'eau est qu'elles doivent être amphiphiles, c'est à dire composées de deux parties: une "tête" hydrophile se trouvant spontanément vers les molécules d'eau et une "queue" hydrophobe qui se dispose avec ses consoeurs.
Parmi les
molécules qui se forment
dans les conditions abiotiques, il en est qui vont posséder
les propriétés adéquates à la formation
de vésicules closes: elles ont été
observées
dans la météorite de Murchison, donc leur synthèse
dans le milieu interstellaire est possible. Elle a été
réalisée expérimentalement (Bernstein, 1999),
à partir de glace d'eau, de ammoniac et de méthanol
exposée à un rayonnement UV. On obtient des
composés
amphiphiles d'une dizaine d'atomes de carbone, qui peuvent s'organiser
en vésicules closes dans l'eau liquide. Outre les UV, les
protons émis dans les vents stellaires peuvent aussi conduire
à la formation de ces molécules. Cela permet leur
synthèse dans les profondeurs sombres et froides (- 258 °C!)
des nuages protoplanétaires, là où ne
pénétrent pas les photons UV (Dworkin, 2004).
Ces molécules pourraient être, par exemple, des phosphates de polyprényl (polymères de l'isoprène) qui peuvent être synthésisés à partir de formaldéhyde et d'acide phosphorique. Ces molécules ne sont pas des inconnues: elles sont présente dans toutes le membranes ou elles jouent le rôle de stabilisateurs (caroténoïdes, cholestérol), de transporteur (ubiquinones) ou de point d'ancrage (géranyl-géraniol, farnésol) pour d'autres molécules.
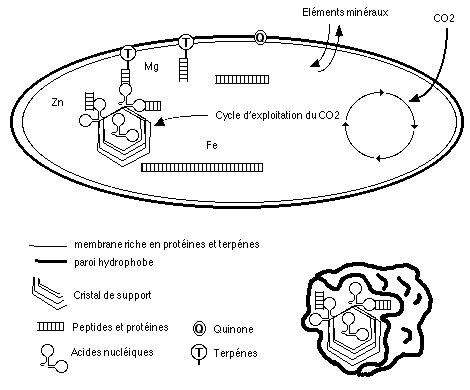 |
Voici a quoi aurait pu ressembler un des premiers organismes vivants: sur un ou plusieurs supports cristallins, les acides nucléiques sont reliés par des peptides à une membrane terpénique. L'évolution moléculaire tend à augmenter la taille et la complexite des peptides, qui stabilisent l'ensemble. La croissance cristalline créée des tensions sur les mmebranes, aboutissant à leur rupture progressive: la reproduction est en route! Ces organismes fixent le carbone atmosphérique via un cycle qui, outre la production d'énergie, alimente les supports cristallins. Ces derniers vont progressivement perdre de l'importance lorque les capacités autoreproductrices de l'ARN (ou de son ancétre) vont se manifester et s'améliorer. Les peptides, qui sont apparu dans un milieu riche en composé minéraux, utilisent ceux cis pour leur topologie: c'est l'origine des cofacteurs minéraux, ainsi que du rôle joué par le Zn dans le repliement des acides nucléiques. A ce stade, ite missa ect.... |
Si la synthèse de ces molécules a été favorisée sur les grains d'origine interstellaire, ceux ci ont du se retrouver englobés dans des vésicules qui les isolait de l'extérieur, et en particulier du milieu aqueux. A l'intérieur de ces paleomembranes on retrouvait un milieu réactionnel hydrophobe, susceptible d'orienter des molécules, de tordre des acides aminés et de favoriser certaines réactions chimiques. Ainsi, les molécules hydrophobes présentent dans l'eau ont tendance à s'accumuler entre les deux feuillets membranaires. On a donc pu avoir liaison entre les peptides s'ancrant et prenant leur origine dans la membrane et ceux se développant à la surface des grains : les membranes aurait été alors fortement "arrimées " aux particules ribonucléiques, constituant ainsi les premières véritables paleocellules. La croissance continue de la membrane peut en outre déclencher des phénomènes de bourgeonnement, de scission en deux vésicules (Luisi, 1998). Comme nous avons vu que nos particules ribosomiales peuvent se retrouver liées à la membrane, le bourgeonnement de celle ci entraîne une scission de ces particules: la reproduction est en marche!
Vu la taille des premières structures
autoreproductibles, leur auto-enfermement dans des vésicules
a du être rapide. Cette petite taille favorisant, en outre,
les liaisons entre membrane et peptides, eux mêmes liés
à lancêtre de l'ARN: on n'était dés
lors plus très loin d'une cellule !
L'organisation
des premiers
génomes
devait ressembler à ce que nous observons actuellement chez
certains
protozoaires: les séquences génétiques
différentes, de petite taille,
étaient présentes sous forme de nombreuses copies
séparées. Lors des divisions accidentelles, puis
organisées, des premières cellules, les
différentes copies étaient réparties au hasard
entre les cellules filles.
Non seulement
les vésicules
bourgeonnaient, mais elles était aussi capables de fusionner,
mélangeant ainsi des matériaux génétiques
divers dans un grand "marché commun" : les premiers génomes n'ont jamais
été stables, et plutot que de parler de cellules
indiduelles il conviendrait mieux de considérer les premiers
êtres vivants comme les membres d'un réseau
génétique d'échanges qui se stabilisera
ultérieurement (Woese, 1998) pour donner les 3
grandes familles actuelles (procaryotes, archeobactéries et
eucaryotes). On en déduit qu'il n'y a jamais eu, en fait,
d'ancêtre commun à toutes les formes de vies actuelles,
mais une "soupe génétique" commune d'où ont
émergé au moins trois lignées distinctes. On passe en fait de l'idée d'un "ancêtre
commun" à celui d'un "ancêtre
communautaire" !
Le schéma
ci contre
résume
ce que nous savons des premiers pas des formes de vies
élaborées
après leur apparition: comme on le voit, leur environnement
(Zahnle, 1996) était loin d'être d'un calme
olympien
!
| STADES
EVOLUTIFS |
EVENEMENTS
MOLECULAIRES ET CELLULAIRES |
Remarques |
Hypothèses
& idées personelles |
|||||||||||||||||||||||||||||||||||||||||||||||||
| SYNTHESE
PREBIOTIQUE |
Synthèse des principaux
éléments structuraux
(acides aminés, bases,
sucres, nucléosides, nucléotides, acides gras,
cofacteurs...)
Il est exclu que
ces molécules se soient formées ensembles (conditions de
synthèse différentes), quelques unes seulement on du se
concentrer sur des minéraux, et dès cette époque
on a eu des aggrégats plurimoléculaires, et pas des
solutions!)
|
synthèses
réalisées en partie dans le milieu interstellaire et
interplanétaire. Apport capital des
micrométéorites |
||||||||||||||||||||||||||||||||||||||||||||||||||
|
Condensation des
éléments structuraux
(oligonucléotides,
oligopeptides, lipides)
Pas d'ARN mais
son ancêtre moléculaire, peut être très
différent de lui.
|
apparition
des membranes (et monde d'ARN) |
|||||||||||||||||||||||||||||||||||||||||||||||||||
| première
ARN réplicase |
trop
tôt ! |
|||||||||||||||||||||||||||||||||||||||||||||||||||
| EVOLUTION
DU PROGENOTE |
Génomes d'ARN Ils n'ont jamais
existé sous forme d'ARN nu: ce dernier a toujours
été relié à des protéines
jouant d'abord un rôle protecteur, puis catalytique : ce "génome" est en fait
l'ancêtre du ribosome !
|
|||||||||||||||||||||||||||||||||||||||||||||||||||
|
Des ribosomes spécifiques
de peptides définissent le code génétique
(tARN primitifs, r ARN,
aminoacyl tRNA synthases)
|
monde
des RNP (Ribo- Nucleo- particules) |
trop tard ! Les RNP sont en fait
antérieurs aux génomes en tant que tel! |
||||||||||||||||||||||||||||||||||||||||||||||||||
|
Appareil de traduction
dépendant de la matrice
(véritables mARN)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||
| Transcription
et réplication de génomes d'ARN bicaténaires
segmentés |
||||||||||||||||||||||||||||||||||||||||||||||||||||
| Génomes
d'ARN recopiés en ADN (ribonucléoside diphosphate reductase, transcriptase reverse) Les enzymes
cités sont le résultat actuel de 3 milliards
d'années d'évolution moléculaire . Ce recopiage
n'en a pas été un, mais une compétition
assurant une meilleure survie aux "génomes" comprenant de plus
en plus d'ADN
|
Début
de notre monde d'ADN. |
|||||||||||||||||||||||||||||||||||||||||||||||||||
|
PROGENOTE
(génome ADN, thymidilate synthétase, géne à
introns, croissance lente, hétérotrophie) Le
progénote devait avoir un matériel
génétique comprenant ARN mélés
à des prtoéines et à des éléments
minéraux, le tout en plusieurs exemplaires redondants et
variables, dispersés dans le premier "cytoplasme". |
||||||||||||||||||||||||||||||||||||||||||||||||||||
| EVOLUTION
DES CELLULES ET DES ORGANISMES |
|
Cette partie de
l'histoire est mieux connue. Toutefois, il y manque les échanges massifs de gènes entre organismes de régnes différents survenant avant le "seuil Darwinien" Depuis le progénote, lien direct vers les nanobes, seuls reliquats actuels de cette époque. |
||||||||||||||||||||||||||||||||||||||||||||||||||
| ci dessus : d'après Biologie
moléculaire de la cellule, Alberts & al., p. 1275, ed 2001. |
||||||||||||||||||||||||||||||||||||||||||||||||||||